How are pholcid spider collectives like a bicycle peloton?
Hugh Trenchard
Rebecca Boyle, for the New Scientist, reported quite nicely one of the essential aspects of the peloton model discussed in my recent paper on protocooperative behavior [1] (free pre-print), which is that the model strips away the deliberate racing strategies and tactics of cyclists, and shows how certain collective peloton formations are dominated by basic physical/physiological principles.
Boyle also notes very well how these basic physical principles drive the changing shape of a peloton -- which was earlier presented in two collaborative papers [2,3] -- but she is circumspect in relation to other key points of my paper and only hints at something potentially more important in it. At the same time the headline emphasizes only the obvious point that weaker cyclists can gain advantage by drafting.
Key elements, not discussed in the New Scientist article, include that my peloton model shows: 1. a mechanism for how groups divide into smaller groups; 2. the threshold between cooperative behavior and predominantly free-riding behavior, and the features of these phases of behavior; and why these factors are significant in an evolutionary sense.
I have a lot of work ahead to show how the principles outlined in my paper may be applied to other biological systems, which is really the Holy Grail of my research. Rebecca Boyle intimates this in referencing fish schools and the comments of James Herbert-Read and Shaun Killen. And while there is voluminous work ahead to show how peloton principles apply to schools of fish and flocks of birds, it is also instructive to consider how the principles apply in systems that are less obviously physically similar to a peloton.
So, here I explore, just a little, how peloton principles apply to a less obvious analogue: social spiders. Unlike the uniformity of movement we see in schools and flocks, spiders tend to move about in different directions and may be separated by considerable distances, and there are few obvious similarities in group shapes and formations, nor is there an obvious energy savings mechanism among spiders, like drafting.
So the question is, how is this:

Fig 1. South American social spiders feeding on katydid
...like this?

Fig. 2. Tour de White Rock men's pro-1,2 criterium (2013)
I suggest there are also deeper similarities. To see how, let's look at some research by Elizabeth Jakob about group dynamics of pholcid spiders [4]. I'm no spider expert, but note that pholcids are not the same spiders as the social spiders in Fig. 1; as I understand it, pholcids are more like the daddy long-legs we commonly see around B.C. I used Figure 1 because it shows a nice collective of spiders*.
For pholcids, groups can be as large as 15, but the most common composition is two sharing a web -- one small and one large spider [4].
Paraphrasing from the Abstract [4] Jakob conducted experiments to examine the role of individual variation in the dynamics of pholcid group formation. Pholcids (specifically H. pluchei) either share webs or live alone, and individuals shift frequently between these conditions. A spider's decision to move is influenced by its size and recent feeding success. Small hungry spiders joining a web already occupied by a larger one were more likely to abandon the web and build their own than small well-fed spiders. The reverse is true for large hungry spiders, which were more likely to stay in a web already occupied than a large well-fed spider.
Jakob explains why, at p. 18:
Hungry small spiders, closer to the bottom of their energetic reserves, may be more sensitive to the negative effects of competition, and thus more willing to take the risk of abandoning a web for potential gain...The behavior of the larger spiders is less obvious: why do well-fed spiders leave groups? One hypthesis is that H. pluchei judge each others' fighting ability by weight. Spiders that are very similar in size fight most intensely (Jakob 1994). Well fed instar 5 spiders have been close enough to the resident's weight that high-intensity potentially dangerous fights were likely.For more background, Jakob reported in [5] that for H. pluchei (quoting from the Abstract):
The main benefit of group living is likely to be the reduction of the cost of web building when spiderlings take advantage of webs built by larger conspecifics.
So this is where we can try some peloton analysis.
In [1-3] we apply an equation that I call the "peloton convergence ratio" (PCR), which is really just a description of the coupled, interactive, relationship between drafting and non-drafting cyclists.
Now we can further explore the notion of the energy savings quantity and what I call the "StarvationCoefficient". If we say that the StarvationCoeff is the equivalent of the drafting coefficient for cyclists, intuitively it doesn't seem to match: the drafting coefficient implies a reduction in output for the drafting cyclist relative to the lead cyclist, and how does this equate to a spider that saves the cost of building its own web by joining one already constructed?
Consider that the drafting coefficient is really a ratio of the relative costs of a leading and drafting cyclist. The cost is higher for the leading cyclist in terms of power output, and lower for the drafting cyclist. The actual energy savings for the drafting cyclist is 1 - d. For example, if the power output for a drafting cyclist is 300W and the power output for the pace-setter is 400W, then the drafting coefficient is 0.75 and the energy savings 25%.
For pholcids (or any organism for that matter), costs are daily energetic demands that prevent the ultimate cost of death by starvation. If we know how long a spider can go without food before death by starvation, and if we know the costs incurred in building a web, we can show the cost of web construction as a ratio of the ultimate cost of starvation.
I calculate the proportionate energy saving to a young pholcid by not building its own web is between 19% (4/21 days) and 23% (5/21 days), and between 6% (4/60 days) and 8% (5/60 days) for an adult. These values represent the energy saved by not building your own web, while the StarvationCoefficients -- the ratio of the reduced time to starvation due to time lost taken to build your own web, to the total cost of living to starvation (1 - energy savings) -- are 0.81, 0.77, 0.94, and 0.92 respectively.
There may be better ways of determining the energy savings benefit of not building a web, but it seems the time saved from imminent starvation is a reasonable quantity by which to develop a model that accounts for the energy savings of not building a web. There may be other factors too that contribute to energy savings or costs, but let's use this StarvationCoeff and the 19% and 8% energy savings and corresponding StarvationCoefficients to develop a model in principle.
Note the PCR and SCR thresholds are inverted. For cyclists the PCR equation denominator reflects the following cyclists' maximum output; the numerator is the required output set by the front cyclist, and so the drafting coefficient reflects the reduction of the required output. For spiders, the denominator is the minimum energy requirement for living. Also, for SCR, the energy savings component implies that the spider's minimum daily food requirement is less than without the energy savings component, so we multiply the energy savings by the minimum requirement in the denominator, and not by the food quantity actually permitted it by the larger spider (in the numerator).
So despite that the PCR and SCR divergence thresholds are inverted, the parameters as between cyclists and spiders are essentially the same in that they both include: 1. an energy quantity determined by a leading agent; 2. an energy savings mechanism and quantity; 3. an energetic limitation for the follower (either a maximum output, as for cyclists, or a minimum food requirement, as for spiders).
Let's see how this works for an adult spider whose StarvationCoeff is 0.92, but which needs say 1.5 flies/ day, but is only allowed 0.5 flies/day by the larger spider.
What happens if either spider is fed before it wanders onto their neighbors' webs? Being fed means that current daily food requirement decreases. Let's say our adult spider is sufficiently well-fed so that for a few days its daily requirement is now only 0.1 flies/day. Say it's still permitted 0.5 flies/day by the dominant larger spider.
Now it becomes easy to find the rate of underfeeding the smaller spider will tolerate before it abandons the web. We simply apply the situation where SCR = 1, which is the point at which the equation states that the spiders will diverge, or the point at which the smaller spider's daily needs are not met, accounting for the energy saved by not building its own web. So with this we can show how hungry the smaller spider will become at its current rate (if 0.5 flies a day is insufficient to meet its daily caloric needs) before it leaves by determining the number of flies/day in the numerator when SCR=1. Let's say our adult smaller spider requires 1 flies/day for sustenance with the same StarvationCoeff of 0.92. How many flies/day will it tolerate before it decides to abandon the web?
In any event, this all seems somewhat promising, though there is more data in Jakob's paper [4] I can test against the framework here, and further work is required generally to develop a more complete model. Also, in terms of protocooperative behavior, a question arises as to how long a spider will go before it relocates, given its willingness to tradeoff the cost of building its own web for receiving less than daily required food needs.
To conclude, there is some promise in adopting an equation similar to the peloton PCR for pholcid coupling behavior. A spider coupling equation, SCR, involves a description of the energetic requirements of coupled spiders that is similar to that of the coupled energetic requirement of cyclists where there is an energy savings quantity. Namely, SCR incorporates: 1. an energy savings quantity; 2. an energetic output that is determined by a "lead" spider, and; 3. an energy quantity limitation for the following spider. These are the essential features of the peloton PCR coupling equation. The SCR equation may allow predictions of food quantities tolerated by pholcids before leaving webs and other related predictions. The apparent application of an SCR equation supports the universality of principles that determine the collective behavior of pelotons.
[1] Trenchard, H. The peloton superorganism and protocooperative behavior. Applied Mathematics and Computation Volume 270, 1 November 2015, pp. 179–192
[2] Trenchard, H., Ratamero, E., Richardson, A., Perc, M. A deceleration model for bicycle peloton dynamics and group sorting Applied Mathematics and Computation Volume 251, 15 January 2015, 24–34
[3] Trenchard, H.,Richardson, A.,Ratamero, E.,Perc, M., Collective behavior and the identification of phases in bicycle pelotons, Physica A: Statistical Mechanics and its Applications Volume 405, 1 July 2014, pp. 92–103
[4] Jakob, E. Individual decisions and group dynamics: why pholcid spiders join and leave groups, Animal Behaviour Volume 68, Issue 1, July 2004, pp.9–20
[5] Jakob, E. Costs and benefits of group living for pholcid spiderlings: losing food, saving silk, Animal Behaviour Volume 41, Issue 4, April 1991, pp. 711–722
* Figure 1 also comes with a creative commons licence.
Image credit (Fig. 1)
South American social spiders feeding on katydid
© Ken Preston-Mafham / Premaphotos Wildlife
Premaphotos Wildlife
Amberstone, 1 Kirland Road, Bodmin, Cornwall
PL30 5JQ United Kingdom
Tel: +44 (0) 1208 78 258
Fax: +44 (0) 1208 72 302
library@premaphotos.co.uk
http://www.premaphotos.com
In [1-3] we apply an equation that I call the "peloton convergence ratio" (PCR), which is really just a description of the coupled, interactive, relationship between drafting and non-drafting cyclists.
PCR = (P_front * d) / MSO_follow
- P_front is the power output of the leading, non-drafting, cyclist, who sets the pace for the group.
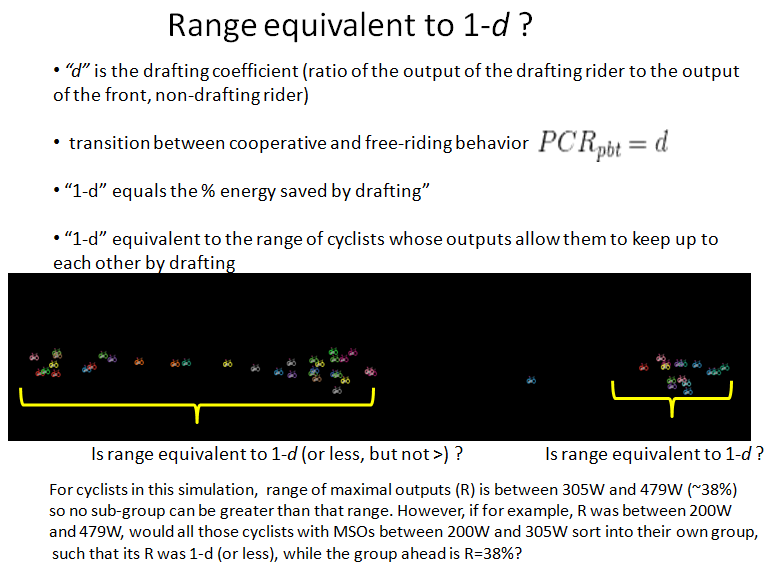
- d is the drafting coefficient (i.e. the ratio of output of the drafting cyclist to the output of the non-drafting cyclist; the actual energy savings is 1-d). It could just as easily be called the "power-output coefficient for coupled cyclists", and is a ratio that describes the relative energetic costs between them, given that a drafting cyclist's output is less than that of the pace-setter. This conceptual subtlety is important when we identify analogues in other systems.
- MSO_follow is the maximum sustainable power output of the following, drafting cyclist.
The MSO_follow of the following rider can vary; it need not be static. In other words, MSO_follow can change according to fatigue or hunger. While we did mention the variability of MSO, in none of [1-3] did we actually model it. However, such a variable MSO is equivalent to changing the speed of the front rider: as the front rider accelerates, the closer that puts the follower to his/her MSO (assuming the follower is seeking to hold the pace of the pacesetter). Similarly, if you reduce the rider's MSO due to sudden fatigue, or the "bonks" due to hunger, he/or she is driven to his/her MSO at a lower speed.
When PCR > 1, coupled cyclists diverge. This simply means that the front cyclist is riding faster than the follower can sustain, even by drafting.
For a small hungry pholcid who decides to abandon the web, we may describe the condition under which it leaves as SCR < 1 (where SCR is "spider convergence ratio"). This is the inverse of the cyclist situation and I will show why, but SCR, like PCR, is still a description of the energetic relationship between the spiders sharing the web, and SCR < 1 describes the threshold at which the smaller pholcid is forced to diverge from that energetic relationship. We can think of this relationship in terms of two pholcids sustaining their individual energetic requirements given that food is distributed between them; and given the smaller spiders' food intake is determined by the larger spider; and in view of energetic costs that are saved by the smaller spider.
In this case, the energy savings is enjoyed by the smaller spider because it saves the cost of constructing its own web by joining a web already constructed by the larger spider. The food energy permitted for the smaller spider's consumption is set by the larger spider, who dominates the web. To understand how these factors might work for spiders and how we might model them, let's first outline some other pholcid behaviors that Jakob identified.
In [5] Jakob observed that (quoting from the Abstract):
...when prey were introduced into group webs, the largest spider in the web that detected the prey won the prey approximately 77% of the time. In spite of this cost, spiderlings were found in groups more often than expected by chance.Also, in [4] Jakob observes that the energy cost of generating silk to make a web is equivalent to about 9 flies, or an average of 4-5 days of foraging. Spiders normally capture 1-2 flies/day. Young pholcids can survive up to 3 weeks without without food (21 days), and adults up to two months [4] (60 days).
Now we can further explore the notion of the energy savings quantity and what I call the "StarvationCoefficient". If we say that the StarvationCoeff is the equivalent of the drafting coefficient for cyclists, intuitively it doesn't seem to match: the drafting coefficient implies a reduction in output for the drafting cyclist relative to the lead cyclist, and how does this equate to a spider that saves the cost of building its own web by joining one already constructed?
Consider that the drafting coefficient is really a ratio of the relative costs of a leading and drafting cyclist. The cost is higher for the leading cyclist in terms of power output, and lower for the drafting cyclist. The actual energy savings for the drafting cyclist is 1 - d. For example, if the power output for a drafting cyclist is 300W and the power output for the pace-setter is 400W, then the drafting coefficient is 0.75 and the energy savings 25%.
For pholcids (or any organism for that matter), costs are daily energetic demands that prevent the ultimate cost of death by starvation. If we know how long a spider can go without food before death by starvation, and if we know the costs incurred in building a web, we can show the cost of web construction as a ratio of the ultimate cost of starvation.
I calculate the proportionate energy saving to a young pholcid by not building its own web is between 19% (4/21 days) and 23% (5/21 days), and between 6% (4/60 days) and 8% (5/60 days) for an adult. These values represent the energy saved by not building your own web, while the StarvationCoefficients -- the ratio of the reduced time to starvation due to time lost taken to build your own web, to the total cost of living to starvation (1 - energy savings) -- are 0.81, 0.77, 0.94, and 0.92 respectively.
There may be better ways of determining the energy savings benefit of not building a web, but it seems the time saved from imminent starvation is a reasonable quantity by which to develop a model that accounts for the energy savings of not building a web. There may be other factors too that contribute to energy savings or costs, but let's use this StarvationCoeff and the 19% and 8% energy savings and corresponding StarvationCoefficients to develop a model in principle.
Now let's consider the PCR equation and how it is equivalent to the pholcid situation. First, PCR describes the coupled relationship between a lead cyclist and a following drafting cyclist. The lead cyclist sets the pace for the follower, while that pace is some proportion of the maximum ability of the follower. For pholcids, it makes sense to consider the larger spider who has constructed its own web as the "lead" spider who dominates the web and dictates what proportion of the total food available goes to the smaller spider. In this way, the larger spider "sets the pace" for the smaller spider in terms of food energy available. As noted, Jakob found the dominant spider won food 77% of the time [5]. So, roughly speaking, if the web catches 2 flies/day, the big spider will get 1.5 spiders, while the small spider gets 0.5 flies/day. This may or may not meet the needs of the small spider. But if 0.5 flies/day is insufficient for its needs, eventually the small spider will reach a threshold point of hunger and will be forced to abandon the shared web in order to make its own.
So for pholcids,
SCR = EnergySet_lead / (EnergyRequired_follower * StarvationCoefficient_follower)
- SCR is "spider convergence ratio".
- EnergySet_lead is the number of flies/day allowed for the smaller spider as determined by the larger spider.
- StarvationCoefficient_follower is the energy cost in days without food as a proportion of the total cost (starvation) in days to starvation, minus the number of days saved by not building a web (or some other energy saving not considered here); i.e. StarvationCoefficient = (1 - energy saved by not building a web).
- EnergyCost_follower is the minimum number of flies required per day for the smaller spider to sustain life.
So for pholcids, let's use energy savings 19% for young ones (StarvationCoeff = 0.81), and 8% for adults (StarvationCoeff = 0.92). So let's say the small pholcid on a web gets 0.5 flies/day, as determined by the large spider, but it actually requires 1 fly/day for sustenance.
SCR = (0.5 flies/day) / (1 fly/day * 0.81)
SCR = 0.62In this case, since SCR < 1, the small pholcid is below its daily energy requirement even as offset by the energy saved by not building its own web, and because SCR < 1 indicates spider divergence, the smaller spider will abandon the shared web to make its own.
Note the PCR and SCR thresholds are inverted. For cyclists the PCR equation denominator reflects the following cyclists' maximum output; the numerator is the required output set by the front cyclist, and so the drafting coefficient reflects the reduction of the required output. For spiders, the denominator is the minimum energy requirement for living. Also, for SCR, the energy savings component implies that the spider's minimum daily food requirement is less than without the energy savings component, so we multiply the energy savings by the minimum requirement in the denominator, and not by the food quantity actually permitted it by the larger spider (in the numerator).
So despite that the PCR and SCR divergence thresholds are inverted, the parameters as between cyclists and spiders are essentially the same in that they both include: 1. an energy quantity determined by a leading agent; 2. an energy savings mechanism and quantity; 3. an energetic limitation for the follower (either a maximum output, as for cyclists, or a minimum food requirement, as for spiders).
Let's see how this works for an adult spider whose StarvationCoeff is 0.92, but which needs say 1.5 flies/ day, but is only allowed 0.5 flies/day by the larger spider.
SCR = (0.5 flies/day) / (1.5 flies/day * 0.92)
SCR = 0.36Here SCR is even smaller than for the example above and, according to this result, this spider will likely have chosen to leave well before it got to this state of hunger.
What happens if either spider is fed before it wanders onto their neighbors' webs? Being fed means that current daily food requirement decreases. Let's say our adult spider is sufficiently well-fed so that for a few days its daily requirement is now only 0.1 flies/day. Say it's still permitted 0.5 flies/day by the dominant larger spider.
SCR = (0.5 flies/day) / (0.1 flies/day * 0.92)
SCR = 5.43Here SCR > 1, and the spider chooses to stay, since its needs are comfortably met. Eventually, as it burns off the food energy from its current state of satiety, the daily requirement will begin to increase again, and the SCR value will fall.
Now it becomes easy to find the rate of underfeeding the smaller spider will tolerate before it abandons the web. We simply apply the situation where SCR = 1, which is the point at which the equation states that the spiders will diverge, or the point at which the smaller spider's daily needs are not met, accounting for the energy saved by not building its own web. So with this we can show how hungry the smaller spider will become at its current rate (if 0.5 flies a day is insufficient to meet its daily caloric needs) before it leaves by determining the number of flies/day in the numerator when SCR=1. Let's say our adult smaller spider requires 1 flies/day for sustenance with the same StarvationCoeff of 0.92. How many flies/day will it tolerate before it decides to abandon the web?
1 = (x flies/day) / ( 1 * 0.92)
x= 0.92Here, even though the spider requires 1 fly/day, it will tolerate 0.92 flies/day -- less than its daily requirement -- for some time because of the energy saved by not having to build its own web. Of course it can't sustain this indefinitely. As the spider becomes hungrier, its daily requirement also increases. At some point it will not tolerate only 0.92 flies/day.
In any event, this all seems somewhat promising, though there is more data in Jakob's paper [4] I can test against the framework here, and further work is required generally to develop a more complete model. Also, in terms of protocooperative behavior, a question arises as to how long a spider will go before it relocates, given its willingness to tradeoff the cost of building its own web for receiving less than daily required food needs.
To conclude, there is some promise in adopting an equation similar to the peloton PCR for pholcid coupling behavior. A spider coupling equation, SCR, involves a description of the energetic requirements of coupled spiders that is similar to that of the coupled energetic requirement of cyclists where there is an energy savings quantity. Namely, SCR incorporates: 1. an energy savings quantity; 2. an energetic output that is determined by a "lead" spider, and; 3. an energy quantity limitation for the following spider. These are the essential features of the peloton PCR coupling equation. The SCR equation may allow predictions of food quantities tolerated by pholcids before leaving webs and other related predictions. The apparent application of an SCR equation supports the universality of principles that determine the collective behavior of pelotons.
References
[1] Trenchard, H. The peloton superorganism and protocooperative behavior. Applied Mathematics and Computation Volume 270, 1 November 2015, pp. 179–192
[2] Trenchard, H., Ratamero, E., Richardson, A., Perc, M. A deceleration model for bicycle peloton dynamics and group sorting Applied Mathematics and Computation Volume 251, 15 January 2015, 24–34
[3] Trenchard, H.,Richardson, A.,Ratamero, E.,Perc, M., Collective behavior and the identification of phases in bicycle pelotons, Physica A: Statistical Mechanics and its Applications Volume 405, 1 July 2014, pp. 92–103
[4] Jakob, E. Individual decisions and group dynamics: why pholcid spiders join and leave groups, Animal Behaviour Volume 68, Issue 1, July 2004, pp.9–20
[5] Jakob, E. Costs and benefits of group living for pholcid spiderlings: losing food, saving silk, Animal Behaviour Volume 41, Issue 4, April 1991, pp. 711–722
* Figure 1 also comes with a creative commons licence.
Image credit (Fig. 1)
South American social spiders feeding on katydid
© Ken Preston-Mafham / Premaphotos Wildlife
Premaphotos Wildlife
Amberstone, 1 Kirland Road, Bodmin, Cornwall
PL30 5JQ United Kingdom
Tel: +44 (0) 1208 78 258
Fax: +44 (0) 1208 72 302
library@premaphotos.co.uk
http://www.premaphotos.com
Figure 2 is from video footage taken by either myself or Ash Richardson, I'm not sure which.