My latest paper The peloton superorganism and protocooperative behavior is published at last. Obviously any published paper requires a lot of work, but for me this one has required the most so far. I began writing this in late 2014, and it was accepted for publication in early August 2015 after the usual process of review and revisions. I would like to think that while this one was more work than my previous papers, it will have the most impact.
I. Protocooperative behavior
Basically the paper argues for a new concept that I call "protocooperative behavior", or PB for short. While the evidence I present is in relation to bicycle pelotons, I suggest that it applies to any biological system in which there is some energy savings mechanism, such as may be found in bird flocks, or fish schools, among others.
Two phases of PB
PB is defined by two phases of behavior: a phase in which cyclists (or other organisms) proceed at a sufficiently low output or speed for cyclists to pass each other and to share the most costly front position(s); a second phase in which cyclists can maintain the speeds of stronger front riders, but cannot pass them. In the low speed phase, the peloton is high density, and in the higher speed phase, the peloton is stretched in single file. At a second threshold, cyclists decouple and diverge.
Phase 1: Low speed, passing and cooperative sharing of costly position
In the low-speed phase, cyclists can pass because to do so is well within their metabolic capacity. To simulate passing, simulated cyclists are programmed to accelerate randomly within a range of speeds up to their maximum capacities. In this low speed phase, cyclists naturally share the most costly high-drag front positions.
Phase 2. High speed, no-passing, free-riding phase
As speeds increase, cyclists' capacity to pass diminishes until a threshold is reached when they cannot pass at all, yet they can still keep pace with stronger riders ahead (the stretched phase). This is possible due to the power output reductions afforded by drafting. In the stretched phase, cyclists are free-riders by physiological necessity, not by choice.
Individual and team strategies need not be modeled
These behaviors -- passing behavior/ sharing the most costly front position, and maintaining speed of stronger rider while being unable to pass -- need not be modeled by strategic probabilities or game theory. The behaviors self-organize from principles of energy savings, cyclists' maximal capacities, their current output, and a deceleration parameter that is triggered when cyclists are effectively driven over their maximal outputs. This is one of the main features that distinguishes this model from more standard models of cooperative behavior; i.e. no strategies are introduced into the model to generate cooperative behavior.
In that vein, team dynamics are not modeled, so circumstances in which teams might dominate the front at low speeds are not considered. This is important because, in natural biological collectives, we are more likely to see random passing based on inherent natural capacities, and less likely to see "teams" motivated by some tactical reason to dominate given positions. We may see something like that in nature, but I suggest such team-like domination would be observed among a comparatively advanced evolutionary stage than the primitive dynamics I am modeling.
The threshold between the phases
There is a clear threshold between a high-density passing phase, and a stretched phase. This threshold is demarcated by the equivalent of the coefficient of drafting. My paper sets out the details of how this works, but simply put, it is a function of the relationship between the coupled outputs of the cyclists, their maximum capacities, and the energy saved by drafting. I illustrate the transition here:
Cyclists sorting into ranges of output capacities
Additionally, cyclists (or any organism) exhibit a heterogeneous range of maximal outputs (i.e. they are not all the same). If the range of outputs is sufficiently broad, when the strongest riders drive peloton speeds to near maximums, the peloton tends to sort into subgroups in which the range of outputs is equivalent to the energy saved by drafting. My paper details why this is so. Below I illustrate the group sorting hypothesis.
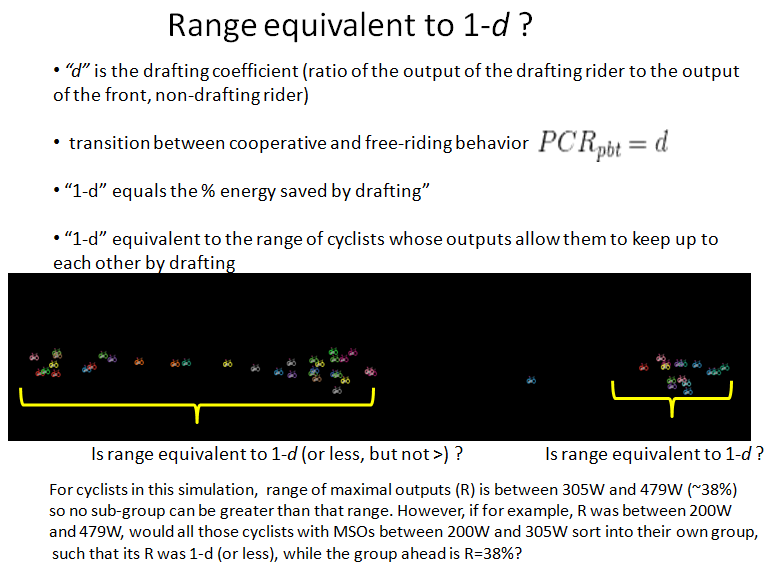
II. Significance
1. The two phases of PB and the threshold between them suggests a primitive form of cooperative behavior that does not rely on any evolutionary strategy (hence "proto" cooperative behavior) -- this proto-behavior may precede other evolutionary mechanisms for cooperation, such as kin selection, reciprocity and others; i.e. it is simply a function of changing speeds and organisms' outputs, coupled by an energy savings mechanism.
2. In its most primitive form, cooperation can't occur unless group members are below a critical threshold of outputs; i.e. they must have some spare energetic resources before they can cooperate, a la the quote from Roosevelt in the paper. So, if resources are strained such that group members are taxed to their physical limits, they will not physically be able to cooperate. This suggests that cooperation evolves in circumstances of "luxury".
As discussed, the critical threshold for "luxury" corresponds to the variation range of maximal outputs that corresponds to the energy savings quantity (1 - d, where d is the energy savings coefficient): when organisms operate below this output threshold, they can cooperate; when above it they can free-ride, but cannot cooperate (Figs 1 and 2), up to a second de-coupling threshold.
So, as group member outputs are increased (and by similar process, resources become scarce), fewer and fewer among the group are within their limits of "luxury" and capable of cooperation; i.e. the strongest cooperate, while the weaker engage in free-riding behavior. Conversely, as outputs fall, or resources become more abundant, cooperation tends to be more widespread, even if it is less necessary for the survival of the group.
I have not researched the literature to find support for this, but it may be seen to be somewhat at odds with some current thinking -- for example, the "ecological constraints hypothesis" suggests that cooperative parenting occurs when resources are scarce [1] rather than when resources are more abundant, as I have suggested. While I need to study the literature a lot more, there may not be an inconsistency at all: what we may be seeing in circumstances of ecological constraint are situations when cooperation is narrowed to the stronger members of the group, while there is an increase in the number of free-riders. So, as a simplistic illustration, the weaker, young members may be fed by the cooperating stronger "parents" for an increased period of time (greater free-riding) before the young are permitted to feed on their own.
3. In some models, such as that of Aviles [2], a cooperation parameter, y, is introduced which, when adjusted, generates different kinds of collective behavior. However, at least insofar as [2] is set out, there is little or no explanation of what criteria are required to adjust the y parameter. The peloton model suggests criteria for tuning that parameter as a function of individual maximal capacities, current outputs, and the energy savings quantity.
4. By modeling a mechanism for group sorting and the resulting range of differences among members of each group, we have a testable mechanism for niche formation, speciation, and group heterogeneity.
Although I didn't say this directly in the paper, here I go so far as to suggest that many branches in the evolutionary tree can be traced to this sorting mechanism. The model suggests an evolutionary mechanism that permits for the shedding of weak group members who are therefore readily susceptible to predators, or who become isolated and lose the opportunity to reproduce. Where whole groups divide and separate, members of each may reproduce, but possibly in very different environments; e.g. one group might "make it over the Himalayas" (thank-you Ross Hooker for the illustration) into the rain-forest, while another may may end up in the high mountains. Indeed it may be possible to show mathematically how the entire range of differences among all existing species conforms to energy savings quantities at critical group sorting points. This would not be an easy task by any means, but it would have to begin with identifying the various energy savings mechanisms enjoyed by different organisms, and the energy savings quantities.
Of course, it is not necessarily true that all biological collectives enjoy energy savings mechanisms. In fact a recent study that shows pigeon flocks involve increased energetic costs due to positional adjustments especially during turning motions [3]*. Still, it is apparent that such mechanisms do appear in many species. Even collections of bacteria tend to move faster than individuals [4] which suggests an energy savings mechanism.
In terms of other applications, there are other types of energy savings mechanisms. For example, any sort of leader-follower situation involves some sort of energy saving for the follower. The person who tramps snow first saves energy for the follower; the person who cuts through the forest makes it easier for those who come after. A teacher saves a student energy because the student does not have to reinvent the wheel, as it were. These are but a few examples. While PB and group sorting for such situations may be more complicated, it is open to consider how the principles I present may be broadly applied.
[1] B.J. Hatchwell and J. Komdeur Ecological constraints, life history traits and the evolution of cooperative breeding.Anim Behav. (2000) 59(6):1079-1086.
[2] Aviles, L. "Cooperation and non-linear dynamics: An ecological perspective on the evolution of sociality." Evolutionary Ecology Research (1999), 1: 459-477
[3] Usherwood, James R., et al. "Flying in a flock comes at a cost in pigeons."Nature 474.7352 (2011): 494-497. [4] Cisneros, Luis H., et al. "Dynamics of swimming bacteria: Transition to directional order at high concentration." Physical Review E 83.6 (2011): 061907. http://www.physics.arizona.edu/~kessler/micro/PhysRevE.83.061907.pdf
* Usherwood et al. seem to focus on energetic costs incurred in banking and turning, as they demonstrate in this video: https://www.youtube.com/watch?v=ssUslfD47l0. I've observed pigeons to fly several km at a stretch in roughly mean straight trajectories, and there are pigeon races over hundreds of km, meaning they are capable of long flight: https://en.wikipedia.org/wiki/Pigeon_racing. I would be interested to see if the same principles of increased costs in flock formations apply in these circumstances.









